主龍類

 Clash Royale CLAN TAG#URR8PPP
Clash Royale CLAN TAG#URR8PPP | 本條目存在隱藏的内容,在一些情况下可能损害讀者的閱覽体验。請協助改善條目,以符合维基百科的标准。(2015年9月12日) |
| 主龍類 化石时期:三疊紀早期到現代 PreЄ Є O S D C P T J K Pg N | |
|---|---|
 | |
| 現存的主龍類包含鱷魚與鳥類 | |
科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 爬行纲 Reptilia |
| 演化支: | 真爬行动物 Eureptilia |
| 演化支: | 卢默龙类 Romeriida |
| 亚纲: | 双孔亚纲 Diapsida |
| 演化支: | 新双弓类 Neodiapsida |
| 演化支: | 蜥类 Sauria |
| 下纲: | 主龙形下纲 Archosauromorpha |
| 演化支: | 主龙类 Archosauria |
| 主要演化支 | |
| |
主龍類(Archosauria)又名初龍類、祖龍類、古龍類,在希臘文意為「具優勢的蜥蜴」,是雙孔亞綱爬行動物的一個主要演化支,包含鱷魚與鳥類,而已經滅亡的恐龍、翼龍目也屬於此。
關於主龍類何時開始出現,有許多爭論。有一派將二疊紀的俄羅斯主龍(Archosaurus rossicus)與原龍(Protorosaurus speneri)視為真正的主龍類,這讓主龍類首次出現於晚二疊紀。而另外一派,將俄羅斯主龍與原龍分類於主龍形類(並非主龍類但關係最近),使得主龍類在早三疊紀奧倫尼克階從主龍形類演化而來。
主龍類主要分為兩個組成部分:鳥頸類和鱷形總目,鱷形總目包括現在還活著的鱷魚以及其他例如勞氏鱷、法索拉鱷、海鱷等已經滅絕的物種;鳥頸類包括恐龍、翼龍和鳥類。
目录
1 譯名
2 特徵
3 主龍類在三疊紀興起
4 主要的種類
5 臀部關節與運動方式
6 滅絕與興盛
7 主龍類的生活方式
7.1 食性
7.2 棲息地
7.3 代謝
7.4 呼吸系統
8 種系發生學
9 参考文献
9.1 引用
9.2 来源
10 外部連結
譯名
主龍類(Archosauria)在希臘文意為「具優勢的蜥蜴」。但許多中文網站與書籍,誤認學名的意思為「古老的蜥蜴」,而翻譯為初龍類、祖龍類、古龍類。
在某些中文網站中,仍然保持早期的傳統分類法,將主龍類(Archosauria)分類成雙孔亞綱的一個下綱,名為主龍下綱、初龍下綱;或是爬行綱的一個亞綱,名為主龍亞綱、初龍亞綱、祖龍亞綱、古龍亞綱。
特徵
主龍類的最基本且最廣為接受的共有衍徵包括:
- 牙齒位在齒槽內,這讓牠們進食時牙齒較不易脫落(這也是槽齒目的名稱來源。不過目前槽齒目被視為早期主龍類的並系群集合,不被現代古生物學家所接受)。
眶前孔(Preorbital fenestrae,位於鼻孔與眼睛之間)減輕了頭顱骨的重量,因為大部分早期主龍類的頭顱骨長而重,這對主龍類是一個非常重要的特徵。眶前孔通常大於眼眶。
下頜孔(Mandibular fenestrae,下頜骨頭上的小洞孔)稍為減輕了下頜的重量。
股骨有第四粗隆部(肌肉附著點)。這個特徵似乎並不重要,卻是恐龍演化出直立姿態的關鍵之一(所有早期恐龍以及許多晚期恐龍是二足動物),也可能與主龍類或牠們的祖先在二疊紀-三疊紀滅絕事件中存活下來有關聯。
主龍類在三疊紀興起

鳥頸類主龍的西里龍、鑲嵌踝類主龍的波羅尼鱷
在二疊紀時期,似哺乳爬行動物是陸地上的優勢脊椎動物,但大部分在二疊紀-三疊紀滅絕事件中滅亡。
草食性的似哺乳爬行動物水龍獸是唯一存活下來的大型陸地動物,並在三疊紀初期成為最繁盛的陸地動物
[1]。
在早三疊紀,主龍類快速地成為陸地上的優勢脊椎動物。關於主龍類為何快速崛起,最常提出的兩個解釋分別為:
- 主龍類演化出直立四肢的過程,比似哺乳爬行動物的演化還快;這讓牠們有更多的耐力,可以在運動時同時呼吸。但這論點是有問題的,因為主龍類擁有優勢地位時,牠們仍處於躺臥或半直立姿勢,類似水龍獸或其他似哺乳爬行動物。
- 早三疊紀的氣候環境普遍乾燥,這是因為當時的陸地集合為盤古大陸。主龍類的保存水分能力,比似哺乳爬行動物好:
- 現代雙孔動物(蜥蜴、蛇、鱷魚、鳥類)都將尿酸以糊狀排洩出來。根據合理的推測,主龍類也將尿酸排泄出來,因此能更好地保存水分。而雙孔動物缺乏皮腺的皮膚也有助於保存水分。
- 現代哺乳類排泄尿素,需要大量的水分以溶解尿素。皮膚也佈有許多皮腺,會流失許多水分。根據推測,似哺乳爬行動物的皮膚擁有類似的特徵,但這仍在爭論中,詳見Palaeos網站。因此牠們在乾燥的環境中擁有劣勢。在澳洲的一個地層顯示出澳洲的上新世至更新世期間,氣候可能一樣乾燥,而該地最大型的陸地掠食動物是古巨蜥(Megalania)與陸地鱷魚,而非哺乳類。
主要的種類
 最原始主龍類 |  鑲嵌踝類主龍 |
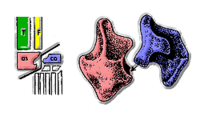 派克鱷與鳥鱷科的反轉型態踝部 |  鳥頸類主龍 |
自從70年代以來,科學家主要依據主龍類的踝部結構來將牠們分類[2]。
- 最早的主龍類擁有原始的中跗骨踝部:距骨與跟骨藉由關節與縫合處固定至脛骨與腓骨。
- 鑲嵌踝類主龍出現於早三疊紀。牠們的距骨藉由關節與縫合處與脛骨連接,而距骨沿者關節嵌入跟骨的窩槽。早期鑲嵌踝類主龍仍然以採取四肢往兩側伸展的步態,但有些晚期鑲嵌踝類主龍發展出完全直立的四肢(尤其是勞氏鱷目)。現代鱷魚也屬於鑲嵌踝類主龍,可依據牠們的行走速度,來採用四肢往兩側伸展或四肢直立的步態行走。
派克鱷以及鳥鱷科擁有與鑲嵌踝類主龍不同的反轉型態踝部,而距骨上有窩槽,而跟骨上有骨樁。- 最早的鳥頸類主龍化石發現於晚三疊紀卡尼階,但目前不清楚鳥頸類主龍如何從鑲嵌踝類主龍演化而來,牠們也可能較早演化出現,也可能從具有原始中跗骨的早期主龍類演化而來。鳥頸類主龍的先進中跗骨形態,是由非常大的距骨與非常小的跟骨所構成,可使踝部關節在單一平面上移動,如同簡易的鉸鏈。這種結構僅適合擁有直立四肢的動物,並在牠們奔跑時提供更多穩定性。鳥頸類主龍與其他主龍類的差別在於:牠們體型較輕巧,通常為小型、牠們的頸部較長,並呈S形彎曲、牠們的頭顱較為輕巧、許多鳥頸類主龍是完全二足動物。鳥頸類主龍與其他主龍類的差別在於:身體通常較小、體格輕、頸部長而呈S形彎曲、頭顱骨較為輕型、而且大部分為二足動物。主龍類的股骨具有第四粗隆部,可使鳥頸類主龍更易演化成二足步態,並提供大腿肌肉更多的力量。在晚三疊紀期間,鳥頸類主龍進一步分化成翼龍目與恐龍[3]。
臀部關節與運動方式

三種不同的臀窩結構與後肢直立方式,從左到右分別為往兩側延展、直立(哺乳類與恐龍)、柱狀直立方式(勞氏鱷目)
如同早期四足總綱動物,早期主龍類的四肢姿勢為往兩側延展。原因為:
- 牠們的臀窩(Hip sockets)朝向兩側。
股骨頂端的頭與股骨本身排列為一直線。
在三疊紀早期到中期,有些主龍類演化支獨自發展出可直立的臀部關節。這個特徵使牠們擁有更多的活動力,而並避免在快速移動時無法同時呼吸。主龍類發展出兩種型態的可直立關節:
- 臀窩朝向兩側,與股骨頭接合。恐龍發展出這種直立方式。
- 臀窩朝向下方,股骨頭與股骨本身平行。不同的主龍類演化支似乎多次個別演化出這種「柱狀直立方式」,例如勞氏鱷目與某些堅蜥目。
滅絕與興盛
鱷目、翼龍目、恐龍、以及離龍目在1億9500萬年前的三疊紀-侏儸紀滅絕事件中存活下來,但其他早期的主龍類則滅絕了,例如植龍目與勞氏鱷目。
恐龍與翼龍目在白堊紀-第三紀滅絕事件中滅亡,但鱷目、離龍目、以及鳥類存活下來。鳥類是主龍類的後代,因此在種系發生學中,鳥類屬於主龍類。離龍目則在漸新世時期滅亡。目前只有鱷目(包括現代鱷魚、長吻鱷、短吻鱷)與鳥類繁盛於現代。
主龍類的生活方式
食性
大部分主龍類是大型掠食動物,但有不同演化支演化成其他的生態位。堅蜥目是草食性動物,有些發展出鱗甲。少數鱷目是草食性動物,例如獅鼻鱷(Simosuchus)、葉齒鱷。早白堊紀的大型腔鱷可能是濾食性動物。蜥腳形亞目、鳥臀目是草食性動物,演化出多樣性的攝食特徵。
棲息地
主龍類主要為陸棲動物,以下例外:
- 鱷目是河流與沼澤裡的優勢動物,甚至進入到海中(真蜥鱷科、地蜥鱷科、森林鱷科)。地蜥鱷科甚至具有類似海豚的外形、槳狀的前肢、錨狀的尾巴、以及平滑無裝甲的皮膚。
- 翼龍目與鳥綱演化出飛行能力,是優勢飛行動物。
代謝
主龍類的代謝仍處於爭議中。牠們的確從冷血動物演化而來,而目前存活的非恐龍主龍類,鱷魚,是冷血動物。但鱷魚擁有一些與溫血動物相關的特徵,因為牠們的促進氧氣供給方式:
- 2個心室與2個心房。哺乳類與鳥類也有2個心室與2個心房。非鱷魚的爬行動物的心臟有3個心房心室,這樣比較沒有效率,因為這樣會將含氧血液與缺氧血液混合在一起,因此會將部份缺氧血液送至身體各處,而非送到肺臟。現代鱷魚雖然擁有四腔室心臟,但與身體相比比例較小,並且與現代哺乳類與鳥類相比,血壓較低。牠們也擁有分導管,可讓牠們位在水面下時,以三腔室心臟運作,以儲存氧氣。
次生顎,可允許動物在進食時可以同時呼吸。
肝瓣,是肺臟的呼吸推動裝置。與哺乳類與鳥類的肺臟推動裝置不同,但根據某些研究人員宣稱,較為類似某些恐龍[4][5]。
有些專家認為鱷魚最初為活耀、溫血的掠食動物,而牠們的主龍類祖先也是溫血動物。研究顯示鱷魚的胚胎具有四腔室心臟,成長後改變為三腔室心臟,以適應水中環境。這些研究人員根據胚胎重演律,牠們提出最初的鱷魚具有四腔室心臟,因此牠們為溫血動物,而後來的鱷魚發展出旁管,重新成為冷血動物,以及水底中的伏擊掠食動物[6][7]。
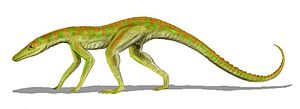
陸鱷是種非常小型、行動敏捷的鱷形類,生存於三疊紀晚期。
如果最初的鱷魚、以及其他的鑲嵌踝類主龍都是溫血動物,這樣可以解決一些演化的謎題:
- 最早期的鱷類,例如陸鱷,是種纖細、長腿的陸地掠食動物,生活方式可能相當活躍,這樣需要非常快速的新陳代謝。而其他的鑲嵌踝類主龍似乎擁有直立的四肢,例如勞氏鱷目。直立的四肢有益於活躍的動物,可讓牠們快速運動時,可以同時呼吸;但不利於緩慢的動物,因為身體站立或趴下時將消耗能量。
- 如果早期主龍類是完全冷血動物,恐龍將以短於似哺乳爬行動物的一半時間,演化為為溫血動物。
呼吸系統
在2010年的一項研究,發現美國短吻鱷有獨特呼吸方式。這項研究發現,當美國短吻鱷吸氣時,吸進的空氣會進入第二支氣管,流經交換氣體的第三支氣管,最後進入第一支氣管。當美國短吻鱷呼氣時,空氣會經由第二支氣管、第三支氣管、第一支氣管,最後從氣管呼出體內[8]。現代鳥類也有類似的呼吸方式,在呼吸過程中,空氣以單向方式在氣囊內流通;而哺乳動物的呼吸方式,則是反覆方向的空氣流通,氣體在肺泡交換。除了現代鳥類以外,許多恐龍演化支也被發現擁有氣囊,可以增進牠們的呼吸效能。
由於現代鱷魚、鳥類都有類似方式的呼吸方式、輔助呼吸系統,有可能是三疊紀的原始主龍類已經演化出這種呼吸系統,而大部分後代都有類似呼吸系統,例如:恐龍、翼龍類、鱷形超目…[8]。研究人員也推測,主龍類的特殊呼吸方式,也是牠們與似哺乳爬行動物的兢爭過程中,逐漸佔優勢的原因之一。在三疊紀時期,大氣層中的含氧量較低,主龍類的特殊呼吸方式,使呼吸效能較高,較具競爭優勢[9][10]。
種系發生學
| 主龍類 Archosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
主龍類 Archosauria
|--鑲嵌踝類主龍 Crurotarsi
| |-?梳棘龍科 Ctenosauriscidae
| `--鱷型踝關節類 Crocodylotarsi
| |--鳥鱷科 Ornithosuchidae
| `--+--植龍目 Phytosauria
| `--Suchia
| |--迅猛鱷科 Prestosuchidae
| `--Rauisuchiformes
| |--堅蜥目 Aetosauria
| `--勞氏鱷目 Rauisuchia
| |--勞氏鱷科 Rauisuchidae
| `--+--Paracrocodylomorpha
| `--鱷形超目 Crocodylomorpha (鱷魚與其近親)
`--鳥頸類主龍 Ornithodira
|--Pterosauromorpha
| |--Scleromochlus
| `--翼龍目 Pterosauria
`--恐龍形態類 Dinosauromorpha
`--恐龍形類 Dinosauriformes
`--恐龍總目 Dinosauria
|--鳥臀目 Ornithischia
`--蜥臀目 Saurischia
`--鳥綱 Aves
参考文献
引用
^ Before the Dinosaurs 互联网档案馆的存檔,存档日期2008-03-15., Discovery Channel
^ Archosauromorpha: Archosauria - Palaeos 互联网档案馆的存檔,存档日期2005-04-05.
^ Archosauromorpha: overview Palaeos 互联网档案馆的存檔,存档日期2005-04-05.
^ Ruben, J.; 等. The metabolic status of some Late Cretaceous dinosaurs. Science. 1996, (273): 120–147. 引文格式1维护:显式使用等标签 (link)
^ Ruben, J.; 等. Lung structure and ventilation in theropod dinosaurs and early birds. Science. 1997, (278): 1267–1247. 引文格式1维护:显式使用等标签 (link)
^ Seymour, R. S., Bennett-Stamper, C. L., Johnston, S. D., Carrier, D. R. and Grigg, G. C. Evidence for endothermic ancestors of crocodiles at the stem of archosaur evolution. Physiol. Biochem. Zool. 2004, 77: 1051–1067.
^ Summers, A.P. Evolution: Warm-hearted crocs. Nature. 2005, 434: 833–834.
^ 8.08.1 Farmer, C. G.; and Sanders, K. Unidirectional airflow in the lungs of alligators. Science. 2010, 327 (5963): 338–340. doi:10.1126/science.1180219. 引文使用过时参数coauthor (帮助)
^ Lisa Grossman. Alligators breath like birds. Science News. January 14, 2010 [January 14, 2010].
^ Oxygen and evolution
来源
.mw-parser-output .refbeginfont-size:90%;margin-bottom:0.5em.mw-parser-output .refbegin-hanging-indents>ullist-style-type:none;margin-left:0.mw-parser-output .refbegin-hanging-indents>ul>li,.mw-parser-output .refbegin-hanging-indents>dl>ddmargin-left:0;padding-left:3.2em;text-indent:-3.2em;list-style:none.mw-parser-output .refbegin-100font-size:100%
- Michael J. Benton (2004), Vertebrate Paleontology, 3rd ed. Blackwell Science Ltd
- Robert L. Carroll (1988), Vertebrate Paleontology and Evolution, W. H. Freeman and Co. New York
外部連結
主龍類各支系加州大學柏克萊分校網站
主龍類 Paleos網站
分類樹 Mikko's Phylogeny Archive網站