白垩纪-第三纪灭绝事件

 Clash Royale CLAN TAG#URR8PPP
Clash Royale CLAN TAG#URR8PPP
| 本条目翻譯品質不佳。(2018年11月17日) |

第三纪灭绝事件-隕石原因想像圖

白垩纪-古近纪界线的暴露位置,位於加拿大亞伯達省德拉姆黑勒鎮附近

白垩纪-古近纪界线的边界,位於美國科羅拉多州的25號州際公路附近。紅箭頭處即為白垩纪-古近纪界线(因富含銥而著名)

來自美國懷俄明州的含白垩纪-古近纪界线岩石。中間的白色黏土層平均銥含量是其他地層的1000倍。
白堊紀-第三紀滅絕事件(簡稱K-T事件、K-T滅絕),又稱為白堊紀﹣古近紀滅絕事件(簡稱K-Pg事件、K-Pg滅絕),俗称恐龙大灭绝,是地球历史上的第四次以及最后一次大規模物种滅絕事件、也是最著名的一次灭绝事件,約發生於6600萬年前[1],中生代白堊紀與新生代第三紀之間,並導致當時地球上的大部分動物與植物消失,包含所有的非鳥類恐龍以外,滄龍科、蛇頸龍目、翼龍目、以及多種的植物與無脊椎動物,也都在這次事件中滅絕。哺乳動物與恐龙的直系后代鳥類則存活下來,並輻射演化,成為新生代的優勢動物[2]。
由於國際地層委員會不再承認第三紀是正式的地質年代名稱,而由古近紀與新近紀取代,因此白堊紀﹣第三紀滅絕事件又可稱為白堊紀﹣古近紀滅絕事件[3]。本中文条目依然采取老式的命名方法。
這個事件因為造成大部份恐龍滅亡與哺乳動物的興起而聞名,但是綜觀地球歷史,二疊紀-三疊紀滅絕事件滅絕了當時地球約90%的生物种类,才是地質年代中最嚴重的生物集體滅絕事件,本次灭绝本不算是最严重的,只是因为以恐龙为代表的生物灭绝而知名度大增而已。
目录
1 判定原则
2 原因
3 滅絕模式
3.1 微生物群
3.2 海生无脊椎動物
3.3 魚類
3.4 陸地無脊椎動物
3.5 陸地植物
3.6 兩棲動物
3.7 非主龍類的爬行動物
3.8 主龍類
3.8.1 鱷形超目
3.8.2 翼龍目
3.8.3 恐龍
3.9 鳥類
3.10 哺乳動物
4 证据
4.1 北美洲化石
4.2 海洋化石
5 經歷時間
6 滅絕原因
6.1 撞擊事件
6.2 德干暗色岩
6.3 多重撞擊事件
6.4 馬斯垂克階的海退事件
7 參考資料
8 外部連結
判定原则
在白堊紀與第三紀的地層之間,有一層富含銥的黏土層,名為白垩纪-古近纪界线。恐龍(不包含鳥類)的化石僅發現於白垩纪-古近纪界线的下層,顯示牠們在這次滅絕事件發生時(或之前)迅速滅絕[4]。有少部份恐龍化石發現於白垩纪-古近纪界线之上,但這些化石被認為是因為侵蝕等作用,而被帶離原本的地點,並沉積在較年輕的沉積層[5][6][7]。
原因
大部分的科學家推測,這次滅絕事件是由一個或多個原因所造成,例如:小行星或彗星引起的撞擊事件、或是長時間的火山爆發。希克蘇魯伯隕石坑等隕石坑以及德干暗色岩的火山爆發,與白垩纪-古近纪界线的時間相近,被認為最有可能與這次滅絕事件的主因。撞擊事件或火山爆發將大量灰塵進入大氣層中,遮闢了陽光,降低了植物的光合作用,進而對全球各地的生態系造成影響。但也有少數科學家認為,這次滅絕事件是緩慢發生的,而滅絕的原因是逐漸改變的海平面與氣候[2]。
滅絕模式
雖然白堊紀﹣第三紀滅絕事件造成許多物種滅絕,但不同的演化支,或是各個演化支內部,呈現出明顯差異的滅絕程度。根據主流學說,大氣層中因撞擊事件或火山爆發所造成的微粒遮蔽了陽光,減少抵達地表的太陽能,導致依賴光合作用的生物衰退或滅絕。如同現今的狀況,在白堊紀晚期,食物鏈底層也是由依賴光合作用的生物構成,例如浮游植物與陸地植物。證據顯示,草食性動物因所依賴的植物衰退,而數量減少;同樣地,頂級掠食者(例如暴龍)也接連受到影響。[8]
顆石藻、軟體動物(包含菊石亞綱、厚殼蛤、水生蝸牛、蚌),還有以上述硬殼動物維生的動物,都在這次滅絕事件中滅亡,或遭受嚴重打擊。例如,被認為以菊石為食的滄龍類,就在此次滅絕事件中滅亡。[9]
雜食性、食蟲性、以及食腐動物在這次滅絕事件中得以存活,可能是因為牠們的食性較多變化性。科學家假設,這些生物以死亡的植物與動物為食,或以生物的有機碎屑為生,因而較不受植物群崩潰滅絕的影響。另外,白堊紀末期似乎沒有完全的草食性或肉食性哺乳動物,牠們和鳥類則藉由食用昆蟲、蚯蚓、蝸牛等小動物而逃過一劫。[2][10][11]
在河流生物群落中,只有少數動物滅亡;因為河流生物群落多以陸地沖刷下來的生物有機碎屑為生,較少直接依賴植物。[12]海洋也有類似的狀況,但較為複雜。生存在浮游帶的動物,所受到的影響遠比生存在海床的動物還大,因為牠們幾乎以活的浮游植物為生。反之,生存在海床的動物,則以生物的有機碎屑為食,或本身可轉換成以生物的有機碎屑為食。[2]
在這次滅絕事件存活下來的生物中,最大型的陸地動物是鱷魚與離龍目,屬半水生動物,並可以生物碎屑為生。現代鱷魚可以食腐為生,並可長達數月未進食;幼年鱷魚的體型小,成長速度慢,在頭幾年多以無脊椎動物、死亡的生物為食。這些特性可能是鱷魚能夠存活過白堊紀末滅絕事件的關鍵。[10]
滅絕事件過後,產生了許多空缺的生態位,生態系統花了長時間才恢復原本的多樣性。[2]
微生物群
在鈣質微型浮游生物的化石記錄中,白堊紀末滅絕事件是最大規模的集體滅絕之一;鈣質微型浮游生物形成的鈣質沉積層,也是白堊紀的名稱來源。這次滅絕事件對鈣質微型浮游生物的影響,只有到種的程度。[13][14]根據統計分析,這段時間的海洋生物物種數量降低,多是因為迅速的滅亡,而非物種形成的減少。[15]由於微生物的化石記錄多由囊胞所構成,而不是所有的甲藻門(Dinoflagellate)都具有囊胞,因此白垩纪-古近纪界线時期的甲藻類化石記錄並不明確,造成相關研究的差異。[2]2004年的一個研究,則認為當時的甲藻類並沒有發生明顯變動。[16]
最早自奧陶紀之後,放射蟲(Radiolaria)一度消失於化石記錄。在白堊紀/第三紀交界,可以找到少量的放射蟲化石。沒有證據顯示當時的放射蟲遭到大規模的滅絕。在古新世早期,放射蟲在南極區相當繁盛,這可能因為寒冷的氣候。[2]接近46%的矽藻物種,繼續存活到上古新世。這顯示白堊紀末的矽藻滅絕程度並不嚴重,層度僅到種的程度。[2][17]
自從1930年代以來,科學家已著手研究白堊紀與第三紀交接時的有孔蟲門(Foraminifera)滅絕狀況。[18][19]
自從撞擊事件可能造成白堊紀末滅絕事件的理論出現後,更多科學家研究白垩纪-古近纪界线時的有孔蟲門滅絕狀況。[2]但科學界對此的意見分為兩派,一派認為有孔蟲門主要因為這次滅絕事件而影響,[20]另一派認為在白堊紀與第三紀交接時期,有孔蟲門有過多次的滅絕與復原。[21][22]
底棲的有孔蟲類主要以生物有機碎屑為食,隨者白堊紀末滅絕事件造成大量海洋生物死亡,眾多的底棲有孔蟲類也跟者滅亡。在海洋生態圈復原後,底棲有孔蟲類的食物來源增加,物種形成也跟着增加。[2]浮游植物在古新世早期復原,提供大型的低棲有孔蟲類食物。直到滅絕事件後的數十萬年,約在古新世早期,低棲生態系統才完全復原。[23][24]
海生无脊椎動物

一個菊石亞綱的化石
化石證據顯示,在白堊紀﹣第三紀滅絕事件的前後,海生無脊椎動物有相當大的變化。但化石紀錄的缺乏,使得無法顯示出真實的滅絕比例。[2]
介形綱(Ostracoda)是甲殼亞門的一綱,這群小型動物在馬斯垂克階上層相當繁盛,在多個地區留下化石紀錄。一個針對介形綱化石的研究,顯示介形綱在古新世時期的多樣性,低於第三紀的其他時期。但目前的研究無法確定介形綱的衰退,是在白堊紀末事件的之前或之後發生。[25][26]
在白堊紀晚期的珊瑚綱石珊瑚目中,有接近60%的屬消失,沒有存活到古新世。近一步的研究顯示,棲息於溫暖、副熱帶、淺海地區的珊瑚,記有98%的種滅亡。而那些棲息於較寒冷、透光帶以下、單獨生長、無法形成珊瑚礁的珊瑚,受到滅絕事件的影響較小。與進行光合作用的海藻共生共存的珊瑚,則在白堊紀﹣第三紀滅絕事件中大量滅亡。[27][28]但除了研究白垩纪-古近纪界线到古新世地層中的珊瑚化石以外,還要參考當時的珊瑚礁生態系統變化。[2]
在白垩纪-古近纪界线之後,頭足綱、棘皮動物、雙殼綱的屬數量,呈現明顯的減少。[2]而大部分的腕足動物,在白堊紀末滅絕事件中存活,並在古新世早期開始多樣化。

一個雙殼綱中的厚殼蛤類化石,發現於阿拉伯聯合大公國的白堊紀末地層。圖中的量尺長10公釐。
在軟體動物頭足綱中,大部分的物種在這次滅絕事件滅亡,只有鸚鵡螺亞綱(以現代的鸚鵡螺為代表)與鞘亞綱(演化成現代的章魚、魷魚、烏賊)存活至今。這些滅亡的物種包含獨特的箭石目,以及高度多樣性、分佈廣泛且數量眾多的菊石。研究人員指出鸚鵡螺亞綱產的卵較大、數量較少,是牠們存活下來的重要原因。而菊石產的卵數量眾多、以浮游生物方式生存,因此在白堊紀末滅絕事件中滅亡。在全球各地的菊石滅亡之後,鸚鵡螺類的外殼開始輻射演化,演化出不同的形狀,構造日益複雜,與昔日的菊石類似。[29][30]
在棘皮動物中,接近35%的屬在這次滅絕事件中消失。其中,生存在低緯度、淺水的棘皮動物滅亡比例最高;而生存在中緯度、深海區域的棘皮動物,被影響的程度則較小。這種滅絕模式的成因可能是棲息地的消失,尤其是淺水地區的碳酸鹽岩台地。[31]
其他的無脊椎動物,例如厚殼蛤類(可形成礁的蛤蜊)與疊瓦蛤(現代扇貝的大型近親),則在白堊紀﹣第三紀滅絕事件中全數滅亡。[32][33]
魚類
在白垩纪-古近纪界线的上下地層,有大量的有頜魚類化石,可供科學家研究這些海生脊椎動物的滅絕模式。在軟骨魚綱中,有接近80%的鯊魚、鰩目、魟魚的科存活過這次滅絕事件;[2]在真骨下綱中,只有少於10%的科消失。[34]在南極洲附近的西摩島,化石紀錄顯示白垩纪-古近纪界线的上層有大量的硬骨魚死亡。科學家推論當時的魚類曾經面臨環境變動,而K-T事件應是這次環境變動的主因。[35]但是,海洋與淡水環境,減輕了魚類所遭受環境變動的影響。[36]
陸地無脊椎動物
在北美洲的14個地點,開花植物化石被用來研究昆蟲的多樣性變化與滅絕比例。研究人員發現,在白垩纪-古近纪界线前的白堊紀地層,有豐富的植物,昆蟲很多樣化。在古新世早期(約滅絕事件後170萬年),植物群仍相當多樣化,來自昆蟲的採食則較少。[37][38]
陸地植物
在白堊紀﹣第三紀滅絕事件發生時,沒有廣佈於全球的植物可供研究。[39]但個別地區的植物群連續狀況可供研究。在北美洲,白堊紀﹣第三紀滅絕事件發生時有大量植物滅亡,但在該次滅絕事件發生前,已有明顯的植物群變化。[40]
在南半球的高緯度地區,例如紐西蘭與南極洲,植物群的物種並沒有發生大量消失,但在植物的繁盛狀況則有短期、劇烈的變化。[37][41]在北美洲,有接近57%的植物物種滅亡。在古新世,化石記錄中的蕨葉,顯示蕨類開始再度興盛,植物群開始復原。在1980年的聖海倫火山爆發後,也出現類似的蕨類復興狀況。[42]
由於白堊紀末滅絕事件,對植物群造成整體性的破壞,當時的腐生生物大量激增,例如真菌,這些生物不需依靠光合作用,而分解死亡植物以吸收營養。真菌的繁盛只持續了數年,當大氣層恢復正常後,光合作用植物(例如蕨類)開始再度生長。[43]
兩棲動物
證據顯示,兩棲動物沒有因為白堊紀﹣第三紀滅絕事件而大量滅亡,大多數成員都存活下來。[2]數個針對蒙大拿州的蠑螈化石研究指出,七個蠑螈屬中有六個屬存活過這次滅絕事件。[44]
青蛙存活到古新世,似乎只有少數種滅亡。但在化石紀錄中,青蛙的科與屬數量並不清楚。[2]一個針對蒙大拿州的三個屬的青蛙研究指出,牠們並沒有受到白堊紀末滅絕事件的影響。[44]這些數據顯示,沒有兩棲類的科在這次滅絕事件中滅亡,或是受到打擊。原因是兩棲類可在水中生存,或者在沉積物、土壤、樹木中築穴、或是在岩層中的洞存活。[36]
非主龍類的爬行動物
在白堊紀﹣第三紀滅絕事件中,非主龍類的爬行動物有龜鱉目、鱗龍超目(包含現今的蛇、蜥蜴、蚓蜥)存活下來,此外還有半水生的離龍目(屬於原始的主龍形下綱,消失於中新世)也倖存過這次滅絕事件。[2]有超過80%的白堊紀烏龜,存活過這次事件。在白堊紀末,總計有六個烏龜的科,都存活到第三紀,並存活至現今。[45]
現存的鱗龍類包含:喙頭蜥目、有鱗目。在中生代早期,喙頭蜥目是群相當成功的動物,而且分佈廣泛,但在白堊紀中期開始衰退。喙頭蜥目目前僅存一屬,僅存於紐西蘭。[46]
現存的有鱗目包含:蜥蜴、蛇、以及蚓蜥,牠們在侏儸紀佔據了許多不同的生態位,並成功地存活到白堊紀。有鱗目存活過白堊紀末滅絕事件,成為現今最成功、最多樣化的爬行動物,目前總計有超過6,000個種。目前未知有任何陸生有鱗目的科,在這次滅絕事件中消失,化石證據也顯示牠們的數量沒有明顯的變化。這些陸生有鱗目動物的體型小、適應性強的代謝率、以及可改變棲息環境的能力,是牠們在白堊紀晚期/古新世早期倖存的原因。[2][45]
非主龍類的海生爬行動物,包含滄龍類、蛇頸龍類,牠們是白堊紀時的優勢海生掠食動物,但在白堊紀末滅亡。[47][48]
主龍類
主龍類包含兩個主要演化支,鑲嵌踝類主龍包含鱷魚與其近親,鳥頸類主龍包含翼龍目、恐龍、以及鳥類。
鱷形超目
在馬斯垂克階的化石紀錄中,鱷形超目有10個科存在者,其中5科在白垩纪-古近纪界线後消失,[49]其餘5科存活到古新世。這些倖存的鱷魚大多棲息於淡水與陸地環境,唯有森林鱷科同時存活於淡水與海洋環境。由此可看出有50%的鱷魚滅絕,而且沒有大型鱷魚存活下來,例如北美洲的恐鱷。[2]鱷魚在滅絕事件後大量存活的原因,可能是牠們的水生生態位與穴居的生活方式,這樣可以減少環境劇烈變動的影響。[36]在2008年,Stéphane Jouve與其同事提出森林鱷科的幼年體生活於淡水環境,如同今日的海生幼年鱷魚,所以當其他海生爬行動物滅亡時,森林鱷科仍得以繼續倖存。淡水環境受到白堊紀﹣第三紀滅絕事件的影響,不若海洋環境來得大。[50]
翼龍目
在馬斯垂克階,唯一確定存在的翼龍類是神龍翼龍科,神龍翼龍科也在滅絕事件中滅亡了。在白堊紀中期,仍有10科大型翼龍類存活者,而後逐漸衰退。較小型的翼龍類則早在馬斯垂克階以前就已滅亡。在馬斯垂克階前,已出現小型動物衰退,而大型動物更為繁盛的現象。在這個時期,新鳥亞綱開始多樣化,並取代其他原始鳥類與翼龍類的生態位,這些動物之間可能出現直接的競爭,或只是新鳥亞綱佔據這些消失物種的空白生態位。[36][51][52]
恐龍

暴龍覓食的重建圖
許多白堊紀末滅絕事件的研究,都關注於恐龍如何滅絕。除了少數的爭議研究,大部分科學家同意恐龍(不包含鳥類)在白堊紀末滅絕事件中滅絕。關於恐龍的滅絕過程分為兩派意見,第一派認為在白堊紀的最後數百萬年,恐龍的多樣性已出現衰退,第二派則認為,在最後數百萬年,並沒有跡象顯示恐龍的衰退。而目前科學家仍無法根據這個時期的恐龍化石,在這兩派意見中做出定論。[53]目前沒有證據顯示馬斯垂克階晚期的恐龍,有穴居、游泳、潛水等習性,所以無法減低環境變動所帶來的影響。某些小型的恐龍可能會存活下來,但无论草食性或肉食性恐龙都會面臨植物或獵物的短缺。[36]
近年來,關於恐龍是內溫性動物的證據日漸增多,恐龍與近親鱷魚的不同代謝程度,有助於科學家研究牠們在白堊紀末的滅絕與存活原因。鱷魚屬於冷血動物,可以連續數個月未進食;而恐龍體型接近的溫血動物,需要更多的食物來源,以維持較快的代謝率。因此當K-T事件發生時,整個食物鏈崩潰,需要大量食物的恐龍滅亡,而鱷魚繼續存活。[8]鳥類與哺乳類等內溫性動物,可能因為體型較小而所需食物較少,加上其他因素,得以在這次滅絕事件倖存。[54]
有數個研究人員主張,恐龍的滅絕是逐漸性的,甚至有恐龍存活到古新世。他們宣稱在海爾河組的岩層中,在白垩纪-古近纪界线的上方1.3公尺處,發現了7种恐龍牙齿化石,意味者在白垩纪-古近纪界线後的4萬年,仍有恐龍存活者。[5]在猶他州聖胡安河的白楊山砂岩層(Ojo Alamo Sandstone),發現了鴨嘴龍類的包括股骨在内的34块骨骼化石,附近有古新世的标准花粉,也得到了古地磁证据的支持。科罗拉多州的阿尼马斯组(Animas Formation)含恐龙部分的时代也可能属于古新世。這個發現顯示該種恐龍存活到第三紀,接近6450萬年前,白垩纪-古近纪界线後的100萬年。如果這個發現屬實,這種鴨嘴龍類將成為倖免存活的演化支。[6]但是有些人研究认为,上述化石可能因為侵蝕作用,而被帶離原本的地點,而在較年輕的沉積層中再度沉積。[7]2004年的一项研究表明在中国南雄盆地发现的鸭嘴龙化石和蛋化石中发现有第三纪的花粉沉积。[55]
鳥類
大部分古生物學家認為鳥類是唯一存活到今日的恐龍。在鳥類中,除了新鳥亞綱以外的原始鳥類在白堊紀末滅絕,包含當時相當繁盛的反鳥亞綱與黃昏鳥目。[56]數個關於鳥類化石的研究顯示,在白垩纪-古近纪界线以前,鳥類的物種已出現多樣化現象,而鴨、雞、以及平胸鳥類的近親已經出現,並與恐龍共同生存。[57]許多新鳥亞綱因為能夠潛水、游泳、或是在水中與沼澤尋找庇護地,因此得以在這次滅絕事件中存活。此外,許多鳥類能夠築穴,或生存在樹洞或白蟻巢中,牠們也存活過這次滅絕事件。這些存活下來的鳥類,在滅絕事件後佔據許多恐龍的原本生態位。[36]
哺乳動物

海爾河組
所有白堊紀的哺乳動物主要族系,包含:卵生的單孔目、多瘤齒獸目、有袋下綱、真獸下綱、磔齒獸超科(Dryolestoidea,原始的獸亞綱)、[58]以及岡瓦那獸亞目(Gondwanatheria,屬於異獸亞綱),[59]都存活過白堊紀﹣第三紀滅絕事件,但都遭受重大損失。生存在北美洲與亞洲的原始有袋類三角齒獸超科(Deltatheroida)遭到絕種。[60]在北美洲的海爾河組,10種多瘤齒獸目中有至少一半成員,以及11種有袋類,都遭到滅亡。[53]
在白堊紀﹣第三紀滅絕事件的前3000萬年,哺乳動物開始多樣化。白堊紀﹣第三紀滅絕事件阻礙了哺乳動物的進一步多樣化。[61]目前的研究指出,儘管恐龍的消失留下大量生態位,但哺乳動物並未因此爆炸性多樣化。[62]一個研究指出,數個哺乳動物的目,在白堊紀﹣第三紀滅絕事件後不久開始多樣化,包含翼手目(蝙蝠)、鯨偶蹄目(鯨魚與海豚、以及偶蹄類);[62]但另一個研究則提出不同看法,認為在白堊紀﹣第三紀滅絕事件後,只有有袋類開始多樣化。[61]
在K-T事件以前,哺乳動物的體型通常很小,接近老鼠的體型,使牠們容易找到庇護地。此外,某些早期單孔目、有袋類、以及真獸類是半水生或穴居動物,如同許多現今哺乳動物的棲息環境。半水生或穴居哺乳動物可在白堊紀﹣第三紀滅絕事件造成的環境壓力中找到庇護所。[36]
证据
北美洲化石

在白堊紀﹣第三紀滅絕事件發生前的地層,有相當豐富的恐龍化石
在北美洲的陸相地層層序中,馬斯垂克階晚期的豐富孢粉型態與白垩纪-古近纪界线後的蕨葉,最能突顯兩個不同時期地層的差異。[39]
在白垩纪-古近纪界线發生前,已知最富含恐龍化石的地層位於北美洲西部,尤其是美國蒙大拿州的海爾河組(年代為馬斯垂克階晚期)。若與海爾河組與加拿大亞伯達省與蒙大拿州的朱迪斯河組/恐龍公園組(年代較早,約7500萬年前)相比較,可得出白堊紀最後1000萬年的恐龍數目變化。這些地層的地理分布有限,僅分布於北美洲的部分地區。[53]
在坎潘階中到晚期的地層中,恐龍的多樣性超過其他時期。馬斯垂克階晚期出現數種大型恐龍:暴龍、甲龍、厚頭龍、三角龍、以及牛角龍,[63]顯示在滅絕事件發生前,這個地區的食物相當豐富。
除了豐富的恐龍化石,植物化石紀錄可重建出植物物種的大幅減少,成為滅絕事件的證據。在白垩纪-古近纪界线以上的地層,花粉化石證實該時期的優勢植物是被子植物;在交界處的地層,花粉化石很少,主要是蕨葉化石。[64]從白垩纪-古近纪界线往上層探索,可以看出花粉化石的數量逐漸增多。在火山爆發後的地區,也可找到這種植物化石的改變模式。[65]
海洋化石
海生浮游生物的大規模、迅速滅亡,發生於白垩纪-古近纪界线時。[66]早在白垩纪-古近纪界线前,菊石亞綱已有小規模、緩慢的衰退,可能與白堊紀晚期的海退有關,其餘的屬在白垩纪-古近纪界线時滅亡。雙殼綱疊瓦蛤科的大部分物種,在白垩纪-古近纪界线前已經逐漸滅亡;同樣在白堊紀末期,菊石的多樣性也出現小規模、逐漸的衰退。[67]研究顯示,在白堊紀晚期的海洋中,已出現數種不同的滅絕模式,發生時間部分交錯,這些滅絕模式結束於白堊紀末滅絕事件。[67]
經歷時間
關於白堊紀﹣第三紀滅絕事件的持續時間,長久以來仍處於爭議中,因為某些理論認為這次滅絕事件的過程很短(從數年到數千年之間),而其他的理論則認為滅絕的過程持續了長時間。由於不可能找到一個物種的完整化石紀錄,所以已滅絕物種的真正滅絕時間,可能晚於最後的化石紀錄。因此,滅絕事件的經歷時間難以測定。[68]科學家已發現少數橫跨白垩纪-古近纪界线、帶有化石的地層,年代從白垩纪-古近纪界线前數百萬年,到白垩纪-古近纪界线後近百萬年。[2]
滅絕原因
關於白堊紀﹣第三紀滅絕事件的成因,科學家們目前已提出數個理論。這些理論多關注於撞擊事件,或是火山爆發,某些理論甚至認為兩者都是原因[69]。在2004年,J. David Archibald與David E. Fastovsky試圖提出一個結合多重原因的滅絕理論,包含:火山爆發、海退、以及撞擊事件。恐龍是當時最大的脊椎動物,首先受到環境改變的衝擊,多樣性開始衰退。火山爆發噴出的懸浮粒子,使得全球氣候逐漸冷卻、乾旱。最後,撞擊事件導致依賴光合作用的食物鏈崩潰,並衝擊已經衰退的陸地食物鏈與海洋食物鏈。多重原因理論與單一原因理論的差別在於,單一原因難以達成大規模的滅絕事件,也難以解釋滅絕的模式。[53]
撞擊事件

小行星撞擊地球的過程想像圖

位於猶加敦半島的希克蘇魯伯隕石坑,直徑達180公里。
在1980年,諾貝爾物理學獎得主路易斯·阿尔瓦雷茨、與兒子沃爾特·阿爾瓦雷茨、弗蘭克·阿薩羅(Frank Asaro)、海倫·米歇爾(Helen Michel)等人發現,全球的白堊紀與第三紀交接地層,地層中的銥含量高於正常標準;舉原始研究中的兩個地層為例,銥含量分別是正常標準的30倍、130倍。銥(Ir)是種地球地殼中非常少見的金屬,屬於親鐵元素;大部分的親鐵元素在行星分化的過程中沉降到地核部分,例如鐵。由於大部分小行星與彗星常發現銥元素,阿爾瓦雷茨等人認為在白堊紀與第三紀的交接時期,曾有顆小行星撞擊地球表面。[70]早在1950年代時,已經有類似的撞擊事件理論,但當時沒有發現任何證據。[71]
在2007年9月,美國科羅拉多州美國西南研究院的研究人員威廉·波特克(William Bottke)與數位捷克籍的科學家,利用電腦計算出希克蘇魯伯撞擊事件的可能肇事者。根據他們的模擬計算結果,該隕石坑有90%的可能性是由一個名為巴普提斯蒂娜的小行星所造成的,該小行星直徑接近160公里,目前運行於火星與木星之間;根據推論,在1億6000萬年前,巴普提斯蒂娜小行星被一顆直徑約55公里的未命名小行星撞擊後粉碎,主要的碎片形成巴普提斯蒂娜族小行星帶,而某些碎片則闖入地球的公轉軌道,其中一顆直徑10公里的碎片,在6,500萬年前撞擊了墨西哥尤卡坦半島,形成希克蘇魯伯隕石坑。另外,月球表面的第谷環形山形成於1億800萬年前,也可能是巴普提斯蒂娜族小行星帶的撞擊結果。[72]
撞擊事件會造成大量的灰塵進入大氣層,遮闢陽光,時間長達1年或1年之內。另外,大量的硫酸以氣溶膠的形式進入平流層中,使抵達地表的日照量降低10%到20%,妨礙植物進行光合作用;平流層內的氣溶膠完全沉降,可能需要10年之內的時間,這將導致植物、浮游植物的大量消失,而在食物鏈上層的草食性動物、掠食性動物也跟着滅亡。而以生物有機碎屑為食的小型生物,受到滅絕事件的影響較小。[54][66]撞擊事件後的數小時內,會有大量的紅外線穿透大氣層,對曝露在外的生物造成傷害。[36]撞擊與再度落下的噴出物,會造成全球性的火風暴。白堊紀晚期的大氣層氧含量高,會助長各地的火勢;而第三紀早期的大氣層氧含量,呈現下降的跡象。如果當時曾發生全球性的火風暴,大氣層中的二氧化碳含量將上升,並在空氣中的灰塵、微粒沉降後,繼續造成短期的溫室效應。在撞擊事件時存活的生物,可能因上述效應而死亡。[73]
撞擊事件可能會造成酸雨,這依撞擊發生地點的地層成分而定。但科學家指出,酸雨造成的影響相對而言較小,而且最多持續約12年。[66]大自然環境會將酸雨稀釋、中和,而且滅絕事件的部分倖存者對酸雨相當敏感,例如青蛙,顯示酸雨並非滅絕事件的重要因素之一。撞擊產生的灰塵、可能還有帶硫的氣溶膠,會在大氣層中慢慢沉降,最多不超過10年,所以只能解釋迅速的滅絕原因。[74]位於墨西哥尤卡坦半島沿岸的希克蘇魯伯隕石坑,是最符合阿爾瓦雷茨理論的隕石坑。在1978年,格倫·彭菲爾德(Glen Penfield)發現並進行希克蘇魯伯隕石坑的鑑定工作。在1990年,其他研究人員完成希克蘇魯伯隕石坑的研究,該隕石坑呈橢圓形,平均直徑為180公里,規模符合阿爾瓦雷茨理論的推測。[75]
從隕石坑的地點與形狀判斷,該小行星撞擊到陸地與海洋交界,除了造成大量灰塵與微粒,還有大型的海嘯。加勒比海與美國東部的數個地層,在當時為陸相地層,卻發現來自海洋的砂;在其他的海相地層,則發現來自陸地的植物碎屑與岩石。撞擊發生處有碳酸鈣地層,這將產生大量的二氧化硫氣溶膠,也會降低地表的日照量,造成酸雨,對植物、浮游生物、具有鈣質外殼的生物造成傷害,例如顆石藻與軟體動物。在2008年8月,一群科學家利用地震學資料,提出該隕石撞擊到的水域,比往常所認定的更深。他們提出這將造成更多的硫酸氣溶膠,嚴重影響氣候,造成更多酸雨。[76]大部分的古動物學家同意在6,500萬年前曾有發生小行星撞擊地球,但撞擊事件是否還有其他因素,仍在研討、探究中。[22][77]
德干暗色岩
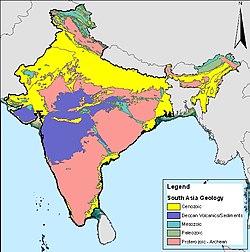
印度的岩石年代圖,粉紅色部分為德干暗色岩爆發形成的火成岩。
在2000年以前,就有德干暗色岩洪流玄武岩是這次滅絕事件的主因,但因為這次火山爆發始於6,800萬年前,並持續超過200萬年,所以當時認為德干暗色岩火山爆發所造成的滅絕事件應該是逐步、緩慢的。但現在有證據顯示,有2/3的德干暗色岩地形,是在一萬年內形成的,發生於6,550萬年前。顯示這次火山爆發造成了快速的滅絕事件,可能在數千年內,但還是遠比撞擊事件長[78][79]。
德干暗色岩火山爆發藉由數種方式造成了滅絕事件,灰塵與二氧化硫與二氧化碳等溫室氣體大量噴出,灰塵使地表的日照量下降,植物的光合作用減少;當灰塵慢慢降落至地表時,火山爆發噴出的二氧化碳氣體,可能增加了溫室效應。許多植物因此而死亡,草食性恐龍因沒有食物而滅亡,而肉食性恐龍也相繼滅絕[79]。在恐龍大規模滅絕前,火山爆發噴出的氣體,似乎與全球暖化有關。有證據指出在希克蘇魯伯撞擊事件之前的50萬年內,大氣溫度曾上升8 °C[78][79]。
在德干暗色岩火山爆發仍被認為是緩慢滅絕事件時,路易斯·阿爾瓦雷茨便已提出古生物學家被少數的資料所混淆。他的警告當時被沒有被接受,但後來針對化石層的研究工作,證明早期的相關研究有誤。目前大部分古生物學家已普遍接受撞擊事件是白堊紀末滅絕事件的主要原因。不過,路易斯·阿爾瓦雷茨的兒子沃爾特·阿爾瓦雷茨(Walter Alvarez)認為在這次撞擊事件前,地球上已發生其他重大事件,例如:海平面下降、造成德干暗色岩的大規模火山爆發,它們可能也是這次滅絕事件的綜合原因之一[80]。
多重撞擊事件
目前已經發現數個隕石坑,形成時間相當於白垩纪-古近纪界线。這些發現指出,當時可能有多次撞擊事件發生;這種模式類似發生在1994年的蘇梅克-列維9號彗星,因為潮汐力而分裂,數個碎片在接下來的幾天內陸續撞擊木星。這些隕石坑包含:烏克蘭的波泰士隕石坑,直徑24公里,形成時間約在距今6517萬年前(誤差值為64萬年)、北海的銀坑隕石坑,直徑20公里,形成時間約在距今6,500萬年前到6,000萬年前。位於印度外海的濕婆隕石坑,當時位於特提斯洋,也是多重撞擊事件的可能隕石坑之一。[81][82][83]
馬斯垂克階的海退事件
有明確的證據顯示,在白堊紀馬斯垂克階發生了中生代以來最大规模的海退。在世界各地的一些馬斯垂克階地層,最早的部分是海床,較早的部分是海岸地層,最晚部分則是陸相地層。這些地層沒有因為造山運動而傾斜、摺曲的跡象,因此最有可能的解釋是海退。目前沒有海退原因的相關證據,較為普遍的解釋是中洋脊的活動降低,而這些巨大海底山脈隨者自身的重量而緩慢沉降於海底地層中。[2][84]
大規模海退造成大陸棚大幅消失,棲息在大陸棚(或稱陸棚、大陸架)的海洋生物最豐富,因此海退可能會造成海洋生物的滅絕。研究也顯示,海平面的變化,不足以造成如此嚴重的菊石滅亡。海平面的下降可能造成大氣環流系統與洋流系統的變化,形成氣候變遷;海洋面積的縮小也會使地表的反照率下降,而使全球氣溫上升。[67]
大規模海退也使許多大陸海消失,例如北美洲的西部內陸海道。這些海域的消失,破壞許多存在於1000萬年之前、生物繁盛的海岸平原,例如恐龍公園組。同時,由於陸地相對上升,河流的長度更長,使淡水的生存區域擴張。海洋與淡水區域的消長變化,使淡水脊椎動物增加,而海洋生物則數量減少,例如鯊魚。[53]
參考資料
^ Renne, Paul R.; Deino, Alan L.; Hilgen, Frederik J.; Kuiper, Klaudia F.; Mark, Darren F.; Mitchell, William S.; Morgan, Leah E.; Mundil, Roland; Smit, Jan. Time Scales of Critical Events Around the Cretaceous-Paleogene Boundary (PDF). Science. 7 February 2013, 339 (6120): 684–687. Bibcode:2013Sci...339..684R. PMID 23393261. doi:10.1126/science.1230492.
^ 2.002.012.022.032.042.052.062.072.082.092.102.112.122.132.142.152.162.172.182.192.20 (英文)MacLeod N, Rawson PF, Forey PL, Banner FT, Boudagher-Fadel MK, Bown PR, Burnett JA, Chambers, P, Culver S, Evans SE, Jeffery C, Kaminski MA, Lord AR, Milner AC, Milner AR, Morris N, Owen E, Rosen BR, Smith AB, Taylor PD, Urquhart E, Young JR. The Cretaceous–Tertiary biotic transition. Journal of the Geological Society. 1997, 154 (2): 265–292. doi:10.1144/gsjgs.154.2.0265. (原始内容存档于2011-02-17).
^ (英文)Fortey R. Life: A Natural History of the First Four Billion Years of Life on Earth. Vintage. 1999: 238–260. ISBN 0375702617.
^ (英文)Fastovsky DE, Sheehan PM. The extinction of the dinosaurs in North America. GSA Today. 2005, 15 (3): 4–10 [2007-05-18]. doi:10.1130/1052-5173(2005)015<4:TEOTDI>2.0.CO;2. (原始内容存档于2011-12-09).
^ 5.05.1 (英文)Sloan RE, Rigby K, Van Valen LM, Gabriel Diane. Gradual dinosaur extinction and simultaneous ungulate radiation in the Hell Creek formation. Science. 1986, 232 (4750): 629–633 [2007-05-18]. PMID 17781415.
^ 6.06.1 (英文)Fassett JE, Lucas SG, Zielinski RA, Budahn JR. Compelling new evidence for Paleocene dinosaurs in the Ojo Alamo Sandstone San Juan Basin, New Mexico and Colorado, USA (PDF). International Conference on Catastrophic Events and Mass Extinctions: Impacts and Beyond, 9-12 July 2000, Vienna, Austria. 2001, 1053: 45–46 [2007-05-18].
^ 7.07.1 (英文)Sullivan RM. No Paleocene dinosaurs in the San Juan Basin, New Mexico. Geological Society of America Abstracts with Programs. 2003, 35 (5): 15 [2007-07-02].
^ 8.08.1 (英文)Wilf P, Johnson KR. Land plant extinction at the end of the Cretaceous: a quantitative analysis of the North Dakota megafloral record. Paleobiology. 2004, 30 (3): 347–368. doi:10.1666/0094-8373(2004)030<0347:LPEATE>2.0.CO;2.
^ (英文)Kauffman E. Mosasaur Predation on Upper Cretaceous Nautiloids and Ammonites from the United States Pacific Coast. Palaios (Society for Sedimentary Geology). 2004, 19 (1): 96–100 [2007-06-17]. doi:10.1669/0883-1351(2004)019<0096:MPOUCN>2.0.CO;2. 引文格式1维护:日期与年 (link)
^ 10.010.1 (英文)Sheehan Peter M, Hansen Thor A. Detritus feeding as a buffer to extinction at the end of the Cretaceous. Geology. 1986, 14 (10): 868–870 [2007-07-04].
^ (英文)Aberhan M, Weidemeyer S, Kieesling W, Scasso RA, Medina FA. Faunal evidence for reduced productivity and uncoordinated recovery in Southern Hemisphere Cretaceous-Paleogene boundary sections. Geology. 2007, 35 (3): 227–230. doi:10.1130/G23197A.1.
^ (英文)Sheehan Peter M, Fastovsky DE. Major extinctions of land-dwelling vertebrates at the Cretaceous–Tertiary boundary, eastern Montana. Geology. 1992, 20 (6): 556–560 [2007-06-22]. doi:10.1130/0091-7613(1992)020<0556:MEOLDV>2.3.CO;2.
^ (英文)Pospichal JJ. Calcareous nannofossils and clastic sediments at the Cretaceous–Tertiary boundary, northeastern Mexico. Geology. 1996, 24 (3): 255–258. doi:10.1130/0091-7613(1996)024<0255:CNACSA>2.3.CO;2.
^ (英文)Bown P. Selective calcareous nannoplankton survivorship at the Cretaceous–Tertiary boundary. Geology. 2005, 33 (8): 653–656.
^ (英文)Bambach RK, Knoll AH, Wang SC. Origination, extinction, and mass depletions of marine diversity. Paleobiology. 2004, 30 (4): 522–542. doi:10.1666/0094-8373(2004)030<0522:OEAMDO>2.0.CO;2.
^ (英文)Gedl P. Dinoflagellate cyst record of the deep-sea Cretaceous-Tertiary boundary at Uzgru, Carpathian Mountains, Czech Republic. Geological Society, London, Special Publications. 2004, 230: 257–273. doi:10.1144/GSL.SP.2004.230.01.13.
^ (英文)MacLeod N. Impacts and marine invertebrate extinctions. Geological Society, London, Special Publications. 1998, 140: 217–246. doi:10.1144/GSL.SP.1998.140.01.16.
^ (英文)Courtillot V. Evolutionary Catastrophes: The Science of Mass Extinction. Cambridge University Press. 1999: 2. ISBN 0521583926.
^ (英文)Glaessner MF. Studien Ober Foraminiferen aus der Kreide und dem Tertiar des Kaukasus. I-Die Foraminiferen der altesten Tertifrschichten des Nordwestkaukasus. Problems in Paleontology. 1937, 2–3: 349–410.
^ (英文)Arenillas I, Arz JA, Molina E & Dupuis C. An Independent Test of Planktic Foraminiferal Turnover across the Cretaceous/Paleogene (K/P) Boundary at El Kef, Tunisia: Catastrophic Mass Extinction and Possible Survivorship. Micropaleontology. 2000, 46 (1): 31–49.
^ (英文)MacLeod N. Nature of the Cretaceous-Tertiary (K-T) planktonic foraminiferal record: stratigraphic confidence intervals, Signor-Lipps effect, and patterns of survivorship. In: Cretaceous–Tertiary Mass Extinctions: Biotic and Environmental Changes (MacLeod N, Keller G, editors). WW Norton. 1996: 85–138. ISBN 0393966572.
^ 22.022.1 (英文)Keller G, Adatte T, Stinnesbeck W, Rebolledo-Vieyra, Fucugauchi JU, Kramar U, Stüben D. Chicxulub impact predates the K-T boundary mass extinction. PNAS. 2004, 101: 3753–3758. PMID 15004276. doi:10.1073/pnas.0400396101.
^ (英文)Galeotti S, Bellagamba M, Kaminski MA, & Montanari A. Deep-sea benthic foraminiferal recolonisation following a volcaniclastic event in the lower Campanian of the Scaglia Rossa Formation (Umbria-Marche Basin, central Italy) (PDF). Marine Micropaleontology. 2002, 44: 57–76 [2007-08-19].
^ (英文)Kuhnt W, Collins ES. 8. Cretaceous to Paleogene benthic foramanifers from the Iberia abyssal plain (PDF). Proceedings of the Ocean Drilling Program, Scientific Results. 1996, 149: 203–216 [2007-08-19].
^ (英文)Coles GP, Ayress MA, and Whatley RC. A comparison of North Atlantic and 20 Pacific deep-sea Ostracoda in Ostracoda and global events (RC Whatley and C Maybury, editors). Chapman & Hall. 1990: 287–305. ISBN 0442311674.
^ (英文)Brouwers EM, De Deckker P. Late Maastrichtian and Danian Ostracode Faunas from Northern Alaska: Reconstructions of Environment and Paleogeography. Palaios. 1993, 8 (2): 140–154. doi:10.2307/3515168.
^ (英文)Vescsei A, Moussavian E. Paleocene reefs on the Maiella Platform Margin, Italy: An example of the effects of the cretaceous/tertiary boundary events on reefs and carbonate platforms. Facies. 1997, 36 (1): 123–139. doi:10.1007/BF02536880.
^ (英文)Rosen BR, Turnsek D. Extinction patterns and biogeography of scleractinian corals across the Cretaceous/Tertiary boundary. Fossil Cnidaria 5.P.. Association of Australasian Paleontology Memoir Number 8, Jell A, Pickett JW (eds). 1989: 355–370.
^ (英文)Ward PD, Kennedy WJ, MacLeod KG, Mount JF. Ammonite and inoceramid bivalve extinction patterns in Cretaceous/Tertiary boundary sections of the Biscay region (southwestern France, northern Spain). Geology. 1991, 19 (12): 1181–1184. doi:10.1130/0091-7613(1991)019<1181:AAIBEP>2.3.CO;2.
^ (英文)Harries PJ, Johnson KR, Cobban WA, Nichols DJ. Marine Cretaceous-Tertiary boundary section in southwestern South Dakota: Comment and Reply. Geology. 2002, 30 (10): 954–955. doi:10.1130/0091-7613(2002)030<0955:MCTBSI>2.0.CO;2.
^ Neraudeau D, Thierry J, Moreau P. Variation in echinoid biodiversity during the Cenomanian–early Turonian transgressive episode in Charentes (France). Bulletin de la Société géologique de France. 1997, 168: 51–61.
^ (英文)Raup DM and Jablonski D. Geography of end-Cretaceous marine bivalve extinctions. Science. 1993, 260 (5110): 971–973. PMID 11537491. doi:10.1126/science.11537491.
^ (英文)MacLeod KG. Extinction of Inoceramid Bivalves in Maastrichtian Strata of the Bay of Biscay Region of France and Spain. Journal of Paleontology. 1994, 68 (5): 1048–1066.
^ (英文)Patterson C. Osteichthyes: Teleostei. In: The Fossil Record 2 (Benton, MJ, editor). Springer. 1993: 621–656. ISBN 0412393808.
^ (英文)Zinsmeister WJ. Discovery of fish mortality horizon at the K-T boundary on Seymour Island: Re-evaluation of events at the end of the Cretaceous. Journal of Paleontology. 1998, 72 (3): 556–571 [2007-08-27].
^ 36.036.136.236.336.436.536.636.7 (英文)Robertson DS, McKenna MC, Toon OB, Hope S, Lillegraven JA. Survival in the first hours of the Cenozoic (PDF). GSA Bulletin. 2004, 116 (5–6): 760–768 [2016-01-08]. doi:10.1130/B25402.1. (原始内容 (PDF)存档于2015-10-22).
^ 37.037.1 (英文)Labandeira Conrad C, Johnson Kirk R, Wilf Peter. Impact of the terminal Cretaceous event on plant–insect associations (PDF). Proceedings of the National Academy of Sciences of the United States of America. 2002, 99 (4): 2061–2066. PMID 11854501. doi:10.1073/pnas.042492999.
^ (英文)Wilf P, Labandeira CC, Johnson KR, Ellis B. Decoupled Plant and Insect Diversity After the End-Cretaceous Extinction. Science. 2006, 313 (5790): 1112–1115. PMID 16931760. doi:10.1126/science.1129569.
^ 39.039.1 (英文)Vajda Vivi, Raine J Ian, Hollis Christopher J. Indication of Global Deforestation at the Cretaceous–Tertiary Boundary by New Zealand Fern Spike. Science. 2001, 294 (5547): 1700–1702. PMID 11721051. doi:10.1126/science.1064706.
^ (英文)Johnson KR, Hickey LJ. Megafloral change across the Cretaceous Tertiary boundary in the northern Great Plains and Rocky Mountains. In: Global Catastrophes in Earth History: An Interdisciplinary Conference on Impacts, Volcanism, and Mass Mortality, Sharpton VI and Ward PD (editors). Geological Society of America. 1991. ISBN 0813722474.
^ (英文)Askin RA, Jacobson SR. Palynological change across the Cretaceous–Tertiary boundary on Seymour Island, Antarctica: environmental and depositional factors. In: Cretaceous–Tertiary Mass Extinctions: Biotic and Environmental Changes, Keller G, MacLeod N (editors). WW Norton. 1996. ISBN 0393966572.
^ (英文)Schultz PH, D'Hondt S. Cretaceous–Tertiary (Chicxulub) impact angle and its consequences. Geology. 1996, 24 (11): 963–967 [2007-07-05].
^ (英文)Vajda V, McLoughlin S. Fungal Proliferation at the Cretaceous–Tertiary Boundary (PDF). Science. 2004, 303: 1489–1490 [2007-07-07]. PMID 15001770. doi:10.1126/science.1093807. (原始内容 (PDF)存档于2007-09-26).
^ 44.044.1 (英文)Archibald JD, Bryant LJ. Differential Cretaceous–Tertiary extinction of nonmarine vertebrates; evidence from northeastern Montana. In: Global Catastrophes in Earth History: an Interdisciplinary Conference on Impacts, Volcanism, and Mass Mortality (Sharpton VL and Ward PD, editors). Geological Society of America, Special Paper. 1990, 247: 549–562.
^ 45.045.1 (英文)Novacek MJ. 100 Million Years of Land Vertebrate Evolution: The Cretaceous-Early Tertiary Transition. Annals of the Missouri Botanical Garden. 1999, 86 (2): 230–258. doi:10.2307/2666178.
^ (英文)Lutz D. Tuatara: A Living Fossil. DIMI Press. 2005. ISBN 0-931625-43-2.
^ (英文)Chatterjee S, Small BJ. New plesiosaurs from the Upper Cretaceous of Antarctica. Geological Society, London, Special Publications. 1989, 47: 197–215 [2007-07-04]. doi:10.1144/GSL.SP.1989.047.01.15.
^ (英文)O'Keefe FR. A cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia). Acta Zoologica Fennica. 2001, 213: 1–63.
^ (英文)Brochu CA. Calibration age and quartet divergence date estimation. Evolution. 2004, 58 (6): 1375–1382. doi:10.1554/03-509.
^ (英文)Jouve, Stéphane; Bardet, Nathalie; Jalil, Nour-Eddine; Suberbiola, Xabier Pereda; Bouya; Baâda; and Amaghzaz, Mbarek. The oldest African crocodylian: phylogeny, paleobiogeography, and differential survivorship of marine reptiles through the Cretaceous-Tertiary Boundary. Journal of Vertebrate Paleontology. 2008, 28 (2): 409–421. 引文使用过时参数coauthors (帮助)
^ (英文)Slack KE, Jones CM, Ando T, Harrison GL, Fordyce RE, Arnason U, Penny D. Early Penguin Fossils, Plus Mitochondrial Genomes, Calibrate Avian Evolution. Molecular Biology and Evolution. 2006, 23: 1144–1155. PMID 16533822. doi:10.1093/molbev/msj124.
^ (英文)Penny D, Phillips MJ. The rise of birds and mammals: are microevolutionary processes sufficient for macroevolution. Trends Ecol Evol. 2004, 19: 516–522. doi:10.1016/j.tree.2004.07.015.
^ 53.053.153.253.353.4 (英文)Archibald J David, Fastovsky David E. Dinosaur Extinction. (编) Weishampel David B, Dodson Peter, Osmólska Halszka (eds.). The Dinosauria 2nd. Berkeley: University of California Press. 2004: 672–684. ISBN 0-520-24209-2.
^ 54.054.1 (英文)Ocampo A, Vajda V, Buffetaut E. Unravelling the Cretaceous–Paleogene (K-T) turnover, evidence from flora, fauna and geology in biological processes associated with impact events (Cockell C, Gilmour I, Koeberl C, editors). SpringerLink. 2006: 197–219. ISBN 3-540-25735-6.
^ (英文)BUCK Brenda J. ; HANSON Andrew D. ; HENGST Richard A. ; SHU-SHENG Hu ;. "Tertiary dinosaurs" in the Nanxiong Basin, Southern China, are reworked from the Cretaceous. The Journal of geology. 2004, 112 (1): 111–118. doi:10.1086/379695.
^ (英文)Hou L, Martin M, Zhou Z, Feduccia A. Early Adaptive Radiation of Birds: Evidence from Fossils from Northeastern China. Science. 1996, 274 (5290): 1164–1167. PMID 8895459. doi:10.1126/science.274.5290.1164.
^ (英文)Clarke JA, Tambussi CP, Noriega JI, Erickson GM, Ketcham RA. Definitive fossil evidence for the extant avian radiation in the Cretaceous. Nature. 2005, 433 (7023): 305–308. doi:10.1038/nature03150.
^ (英文)Gelfo JN & Pascual R. Peligrotherium tropicalis (Mammalia, Dryolestida) from the early Paleocene of Patagonia, a survival from a Mesozoic Gondwanan radiation. Geodiversitas. 2001, 23: 369–379.
^ (英文)Goin FJ, Reguero MA, Pascual R, von Koenigswald W, Woodburne MO, Case JA, Marenssi SA, Vieytes C, Vizcaíno SF. First gondwanatherian mammal from Antarctica. Geological Society, London, Special Publications. 2006, 258: 135–144.
^ (英文)McKenna, Malcolm C.; Bell, Susan K. Classification of Mammals: Above the Species Level. Columbia University Press. 1997. ISBN 978-0-231-11012-9. 引文使用过时参数coauthors (帮助)
^ 61.061.1 Bininda-Emonds ORP, Cardillo M, Jones KE, MacPhee RDE, Beck RMD, Grenyer R, Price SA, Vos RA, Gittleman JLY, Purvis A. The delayed rise of present-day mammals (PDF). Nature. 2007, 446: 507–512 [2008-08-16]. doi:10.1038/nature05634. (原始内容 (PDF)存档于2008-12-17).
^ 62.062.1 (英文)Springer MS, Murphy WJ, Eizirik E, O'Brien SJ. Placental mammal diversification and the Cretaceous–Tertiary boundary (PDF). PNAS. 2003, 100 (3): 1056–1061. PMID 12552136. doi:10.1073/pnas.0334222100.
^ (英文)Dodson Peter. The Horned Dinosaurs: A Natural History. Princeton: Princeton University Press. 1996: 279–281. ISBN 0-691-05900-4.
^ (英文)Online guide to the continental Cretaceous–Tertiary boundary in the Raton basin, Colorado and New Mexico. U.S. Geological Survey. 2004 [2007-07-08]. (原始内容存档于2012-09-11).
^ (英文)Smathers GA, Mueller-Dombois D. Invasion and Recovery of Vegetation after a Volcanic Eruption in Hawaii, Scientific Monograph Number 5. United States National Park Service. 1974 [2007-07-09]. (原始内容存档于2014-04-03).
^ 66.066.166.2 (英文)Pope KO, D'Hondt SL, Marshall CR. Meteorite impact and the mass extinction of species at the Cretaceous/Tertiary boundary. PNAS. 1998, 95 (19): 11028–11029. PMID 9736679. doi:10.1073/pnas.95.19.11028.
^ 67.067.167.2 (英文)Marshall CR, Ward PD. Sudden and Gradual Molluscan Extinctions in the Latest Cretaceous of Western European Tethys. Science. 1996, 274 (5291): 1360–1363. PMID 8910273. doi:10.1126/science.274.5291.1360.
^ (英文)Signor III PW, Lipps JH. Sampling bias, gradual extinction patterns, and catastrophes in the fossil record. In: Geological implications of impacts of large asteroids and comets on the Earth, Silver LT, Schultz PH (editors). Geological Society of America Special Publication. 1982, 190: 291–296.
^ 恐龙之死 可能曾经有三次小行星撞击发生在地球史上大约同一时刻,引发了一个历时久远的超级火山喷发大灾难。
^ (英文)Alvarez LW, Alvarez W, Asaro F, Michel HV. Extraterrestrial cause for the Cretaceous–Tertiary extinction. Science. 1980, 208 (4448): 1095–1108. PMID 17783054. doi:10.1126/science.208.4448.1095.
^ (英文)De Laubenfels MW. Dinosaur extinction: One more hypothesis. Journal of Paleontology. 1956, 30 (1): 207–218.
^ (英文) P, Claeys; Goderis, S. Solar System: Lethal billiards. Nature. 2007-09-05, 449: 30–31. doi:10.1038/449030a. 引文使用过时参数coauthors (帮助); 使用|accessdate=需要含有|url=(帮助)
^ (英文)Pope KO, Baines KH, Ocampo AC, Ivanov BA. Energy, volatile production, and climatic effects of the Chicxulub Cretaceous/Tertiary impact. Journal of Geophysical Research. 1997, 102 (E9): 21645–21664. doi:10.1029/97JE01743. (原始内容存档于2008-04-29).
^ (英文)Kring DA. Environmental consequences of impact cratering events as a function of ambient conditions on Earth. Astrobiology. 2003, 3 (1): 133–152. PMID 12809133. doi:10.1089/153110703321632471.
^ (英文)Pope KO, Ocampo AC, Kinsland GL, Smith R. Surface expression of the Chicxulub crater. Geology. 1996, 24 (6): 527–530. PMID 11539331. doi:10.1130/0091-7613(1996)024<0527:SEOTCC>2.3.CO;2.
^ Marc Airhart. Seismic Images Show Dinosaur-Killing Meteor Made Bigger Splash. Jackson School of Geosciences, The University of Texas at Austin. 2008-01-17. (原始内容存档于2008-02-09) (英语).
^ (英文)Morgan J, Lana C, Kersley A, Coles B, Belcher C, Montanari S, Diaz-Martinez E, Barbosa A, Neumann V. Analyses of shocked quartz at the global K-P boundary indicate an origin from a single, high-angle, oblique impact at Chicxulub. Earth and Planetary Science Letters. 2006, 251 (3–4): 264–279. doi:10.1016/j.epsl.2006.09.009.
^ 78.078.1 (英文)Hofman, C, Féraud, G & Courtillot, V. 40Ar/39Ar dating of mineral separates and whole rocks from the Western Ghats lava pile: further constraints on duration and age of the Deccan traps. Earth and Planetary Science Letters. 2000, 180: 13–27. doi:10.1016/S0012-821X(00)00159-X.
^ 79.079.179.2 (英文)Duncan, RA & Pyle, DG. Rapid eruption of the Deccan flood basalts at the Cretaceous/Tertiary boundary. Nature. 1988, 333: 841–843. doi:10.1038/333841a0.
^ (英文)Alvarez, W. T. rex and the Crater of Doom. Princeton University Press. 1997: 130–146. ISBN 978-0691016306.
^ Mullen L. Debating the Dinosaur Extinction. Astrobiology Magazine. 2004-10-13 [2007-07-11]. (原始内容存档于2009-01-05). (英文)
^ (英文)Mullen L. Multiple impacts. Astrobiology Magazine. 2004-10-20 [2007-07-11]. (原始内容存档于2009-01-05).
^ (英文)Mullen L. Shiva: Another K–T impact?. Astrobiology Magazine. 2004-11-03 [2007-07-11]. (原始内容存档于2009-02-01).
^ (英文)Liangquan L, Keller G. Abrupt deep-sea warming at the end of the Cretaceous. Geology. 1998, 26: 995–998. doi:10.1130/0091-7613(1998)026<0995:ADSWAT>2.3.CO;2.
(英文)Fortey R. Earth: An Intimate History. Vintage. 2005. ISBN 0375706202.
外部連結
- 專門性:
(英文)希克蘇魯伯隕石坑的爭議(普林斯頓大學網站)
(英文)德干暗色岩火山爆發引起的溫室效應(佛蒙特大學網站)
(英文)希克蘇魯伯隕石坑(NASA)
(英文)白堊紀﹣第三紀滅絕事件(加州大學動物學博物館)
(英文)誰殺死了恐龍?(加州大學動物學博物館)
- 新聞性:
(英文)BBC新聞:恐龍的滅絕原因是?科學家懷疑除了隕石撞擊以外,還有其他的元兇。
(英文)國家地理頻道:殺死恐龍的元兇共有四個:氣候變化、火山爆發、希克蘇魯伯隕石坑、以及另一個年代較晚的不明撞擊事件。
白堊紀-第三紀滅絕事件 | |
疑似K-T事件的隕石坑 | |
| 波泰士隕石坑 | 希克蘇魯伯隕石坑 |
| 濕婆隕石坑 | 銀坑隕石坑 |
生物集群灭绝事件 |
|---|
規模稍小的滅絕事件 ↓埃迪卡拉纪末期? ↓劳阶 ↓托阿尔阶 ↓阿普第阶 ↓森諾曼期-土侖期 ↓中新世 ↓石炭纪雨林崩溃 ↓波托米阶末期 ↓寒武纪-奥陶纪 ↓奧陶紀-志留紀 ↓泥盆紀後期 ↓二疊紀-三疊紀 ↓三疊紀-侏羅紀 ↓白堊紀-第三紀 ↓全新世、第四紀 大型滅絕事件 埃迪卡拉纪 侏羅紀 古近纪 新元古代 古生代 中生代 新生代 │ −600 │ −550 │ −500 │ −450 │ −400 │ −350 │ −300 │ −250 │ −200 │ −150 │ −100 │ −50 │ 0 距今百萬年前 |
| |||||||||||||||||||||||||||||||||||||||||||

