三磷酸腺苷合酶

 Clash Royale CLAN TAG#URR8PPP
Clash Royale CLAN TAG#URR8PPP
| ATP合酶 | |||||||
|---|---|---|---|---|---|---|---|
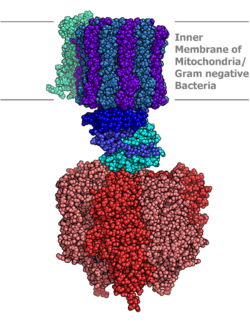 | |||||||
| 通过X射线晶体学测定ATP合酶的分子模型 | |||||||
| |||||||
| 识别码 | |||||||
EC编号 | 3.6.3.14 | ||||||
CAS号 | 9000-83-3 | ||||||
| 数据库 | |||||||
IntEnz | IntEnz浏览 | ||||||
BRENDA | BRENDA入口 | ||||||
ExPASy | NiceZyme浏览 | ||||||
KEGG | KEGG入口 | ||||||
MetaCyc | 代谢路径 | ||||||
PRIAM | 概述 | ||||||
PDB | RCSB PDB PDBj PDBe PDBsum | ||||||
基因本体 | AmiGO / EGO | ||||||
| |||||||
三磷酸腺苷合酶或ATP合酶,三磷酸腺苷酶(ATPase)的一种,在这里并特指F类的FoF1ATP合酶(F Type FoF1 ATP Synthase)。它利用呼吸链产生的质子的电化学势能,通过改变蛋白质的结构来进行三磷酸腺苷(ATP)的合成。ATP是大多数生物体中细胞最常用的“能量通货”。 它由二磷酸腺苷(ADP)和无机磷酸盐(Pi)形成。 ATP合酶催化的总体反应为:
- ADP + Pi + H+out ⇌ ATP + H2O + H+in
ATP合酶由两个主要的亚基Fo和F1组成,它们具有允许ATP产生的旋转运动机制[1][2]。
目录
1 分布
2 位置
3 结构与功能
3.1 F1单元
3.2 FO单元
4 ATP合酶的反应
5 结合模型
6 历史
7 參見
8 參考資料
分布
通过使用电子显微镜,蘑菇状的F型ATP酶可以在真核细胞的粒線体内膜和原核生物的細胞膜上观察到。
位置
在真核细胞中,ATP合酶存在于线粒体的内膜或,Fo亚单位存在于膜内,F1
- F型ATP酶(ATPase)– 也称为‘Phosphorylation Factor’存在在各种生物中,利用电化学势进行ATP的合成。
- P型ATP酶(也称 E1-E2 ATP酶)– 存在在细菌,和真核细胞中,消费ATP进行离子运输。
- V型ATP酶 – 存在在液泡(Vesicle),如高尔基体,溶酶体上,消费ATP进行离子运输
- A型ATP酶 – 存在在古细菌(Archaea)中,虽然有F型ATP酶类似的功能——ATP合成,但是在结构上其更接近与V型ATP酶,反映了古细菌为适应极端条件的进化。
结构与功能
当前,原核生物的F型ATP酶的结构已经比较清楚了:
- F1单元 – α(3个)、β(3个)、γ(1个)、δ(1个)、ε(1个)
- Fo单元 – a(1个)、b(2个)、c(9-12个)
真核生物的F型ATP酶F1单元的种类的数量与原核生物相同、Fo单元的结构与原核生物相似,但是亚单位的数量不是很明瞭。
F1单元
ATP合酶的F1部分是亲水性的,并且负责水解ATP。 子单元αdisplaystyle alpha 

| 子单元 | 人类基因 |
|---|---|
| alpha | ATP5A1, ATPAF2 |
| beta | ATP5B, ATPAF1, C16orf7 |
| gamma | ATP5C1 |
| delta | ATP5D |
| epsilon | ATP5E |
FO单元

FO子单元F6来自ATP合酶的外周茎区。[3]
FO是具有八个子单元的和跨膜环的水不溶性蛋白质。
| 子单元 | 人类基因 |
|---|---|
| a | ATP6 |
| b | ATP5F1 |
| c | ATP5G1, ATP5G2, ATP5G3 |
ATP合酶的反应
F1 单元催化以下ATP合成反应。
ATP ⟵→displaystyle overrightarrow longleftarrowADP+Pi(磷酸)
F1单元催化的反映是可逆的,在进行离子运输时,进行ATP水解反应。
Fo单元形成离子通道,质子可以从中通过:
- H+in ⟵→displaystyle overrightarrow longleftarrow
当质子利用电化学势能通过Fo单元时,可以带动和其连接的轴(γ亚单位),改变F1单元的结构,进而调节F1单元与ATP和ADP:Pi的结合能(Binding Energy),降低ATP生成的活化能,达到ATP合成的目的:
- ADP + Pi + 3 H+out → ATP + 3 H+in

ATP合成的机制:ATP为红色,ADP和磷酸为粉色,旋转的γ亚基为黑色
结合模型
在ATP酶的酶学模型中,验证其γ轴是否旋转占有重要地位,1997年,英国自然杂志(vol. 386, pp. 299–302)刊了日本科学家题为"Direct observation of the rotation of F1-ATPase"的文章,报道了ATP合成酵素F1单元可以通过水解ATP造成γ亚单位(轴)的旋转,并进行了单分子观察和录像、该論文证明了保罗·博耶的「ATP合酶的旋转理论」。同年保罗·博耶,约翰·沃克和延斯·克里斯蒂安·斯科因ATP合酶的研究获得诺贝尔化学奖。
历史
參見
- 電子傳遞鏈
- 叶绿体
- 黄素蛋白
- 線粒體
- 氧化磷酸化
- ATP酶
參考資料
^ Okuno D, Iino R, Noji H. Rotation and structure of FOF1-ATP synthase. Journal of Biochemistry. April 2011, 149 (6): 655–64. PMID 21524994. doi:10.1093/jb/mvr049.
^ Junge W, Nelson N. ATP synthase. Annual Review of Biochemistry. June 2015, 84: 631–57. PMID 25839341. doi:10.1146/annurev-biochem-060614-034124.
^ PDB 1VZS; Carbajo RJ, Silvester JA, Runswick MJ, Walker JE, Neuhaus D. Solution structure of subunit F(6) from the peripheral stalk region of ATP synthase from bovine heart mitochondria. Journal of Molecular Biology. 2004, 342 (2): 593–603. PMID 15327958. doi:10.1016/j.jmb.2004.07.013.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
